![]()
ATTELAGE : CONSEILS QUE LE VÉTÉRINAIRE DOIT
POUVOIR DONNER A TOUT MENEUR
par Dr Guy SOUFFLEUX
De l'énergie du soleil à la force du
cheval (première publication 21 décembre 2007,
dernière mise à jour 21 décembre 2007)
Le soleil, grâce à des réactions
nucléaires, libère de l'énergie qui arrive sur terre sous forme de radiations
électromagnétiques. Une petite quantité de ces radiations est absorbée par
les molécules de chlorophylle des plantes vertes et des algues et va permettre
la synthèse de glucose à partir de l'eau et de dioxyde de carbone. Ce glucose
va permettre à la plante de synthétiser des glucides, des lipides, des
protéines pour assurer sa croissance et sa reproduction.
Le cheval, en consommant les plantes,
va libérer l'énergie des radiations "électromagnétiques qu'elles
avaient emmagasinée, et la transformer en énergie cinétique lorsqu'il est
attelé.
Quelques rappels de physique
énergétique s'imposent pour bien comprendre le sens des réactions
biochimiques qui vont permettre cette conversion d'énergie.
1) L' ATOME
Au niveau atomique, les électrons qui vont s'échanger pour produire des liaisons covalentes entre les atomes, ou qui vont constituer des dipôles moléculaires en se rassemblant plus à une extrémité de la molécule et vont produire des liaisons hydrogène, sont de la plus haute importance.
Un atome est constitué d'un noyau dont les charges électriques positives portées par les protons sont égales aux charges électriques négatives portées par les électrons qui tournent autour. La force attractive entre les charges positives et les charges négatives est la force de Coulomb
F = K q 1 q 2 / D2
On voit tout de suite qu'un noyau ayant beaucoup de protons exerce une force attractive plus grande sur chaque électron, et de ce fait le diamètre des atomes qu'ils soient lourds ou légers est à peu près le même. Ainsi l'uranium a un diamètre 3 fois plus grand que celui de l'hydrogène alors qu'il est 238 fois plus lourd.
Les électrons tournent
autour du noyau dans des orbitales (volume dans lequel on a 90% de chance de
rencontrer l'électron) dans lesquelles la force centrifuge donnée par la
vitesse de rotation équilibre la force de Coulomb. L'électron le plus interne
est soumis à la force la plus forte. Sa vitesse de rotation est la plus
élevée, il a donc l'énergie cinétique la plus forte. A l'inverse,
l'électron le plus externe a l'énergie cinétique la plus faible.
Selon le 1er principe de la
thermodynamique qui indique que dans un système isolé il y a conservation de
l'énergie, lorsqu'un électron s'éloigne du noyau, son énergie cinétique
diminue, donc son énergie potentielle augmente. L'énergie potentielle est
l'énergie contenue dans une particule du fait de sa structure ou du fait de sa
position par rapport à d'autres particules.
L'électron de H (hydrogène), à l'état fondamental (repos) a une énergie de - 13,6 eV, à l'état excité par un photon ultra violet de 10,2 eV il s'éloigne du noyau et son niveau énergétique devient - 3,4 eV. Son énergie potentielle s'est donc accrue de 10,2 eV
L'oxygène est un petit atome pour
lequel il manque 2 électrons pour saturer la 2e orbitale . La force de Coulomb
étant élevée du fait de la courte distance qui sépare le nuage électronique
du noyau de l'atome, il attire les électrons des atomes voisins et est très
réactif.
2) LES REACTIONS CHIMIQUES
Elles répondent au 2e principe de la thermodynamique et à la notion d'entropie. On définit l'entropie d'un système comme sa faculté à subir un changement spontané entre un état et un autre. Ainsi l'entropie d'un système augmente quand il perd de la chaleur, elle diminue quand il en reçoit et elle reste inchangée par un travail effectué en absence de frottement.
L'augmentation de l'entropie va avec l'augmentation du désordre. Ainsi dans le système ordonné renfermant montagnes et collines, l'érosion liée à la gravité aplanit tout, il y disparition de l'énergie potentielle, mélange de toutes les roches devenues sables donc accroissement du désordre, et augmentation de l'entropie. L'entropie sera maximale lorsque tout sera aplani, et le système ne pourra plus évoluer, il aura perdu toute son énergie potentielle. L'entropie définit la direction du temps vers le futur, vers le désordre maximum.
Les réactions chimiques spontanées se font au dépend de l'énergie libre du système (énergie potentielle, énergie cinétique, énergie chimique), elles libèrent de la chaleur et augmentent l'entropie du système suivant l'équation :
delta G = delta H - T delta S
T = température absolue
delta S = variation d'entropie
delta H = production de chaleur = enthalpie
delta G = variation de l'énergie libre du système
A température constante, si la
réaction chimique se fait sans production de chaleur ( H = 0), la diminution de
l'énergie libre ( G est négative dans ce cas) entraîne une augmentation de
l'entropie ( S est positive). Par contre si l'énergie libre augmente,
l'entropie devient négative donc diminue, ce qui est impossible à partir d'une
réaction chimique spontanée en respectant le 2e principe de thermodynamique.
La possible diminution de l'énergie
libre du système est une force qui pousse les réactions chimiques à avoir
lieu.
Une réaction chimique est irréversible lorsque l'énergie libre des réactifs est supérieure à l'énergie libre des produits. Entre les deux termes il y a eu libération de chaleur et augmentation de l'entropie. C'est une réaction exergonique.
2 H2 + O2 => 2 H2 O + chaleur
Pour rendre cette réaction réversible, il faut apporter de l'énergie au système, cela peut-être fait avec des radiations lumineuses. C'est une réaction endergonique.
2 H2 O + photon => 2 H2 + O2
Dans une cellule, lorsqu'une réaction nécessite un apport d'énergie extérieure, celle-ci est fournie par une réaction couplée qui va augmenter l'énergie libre des 1ers réactifs et rendre possible leur réaction.
La réaction couplée la plus fréquente est liée à l'ATP (Adénosine Tri Phosphate)
ATP => ADP + Pi par hydrolyse
Cette réaction libère de l'ADP (Adénosine Di Phosphate) et un groupement phosphate inorganique qui va se combiner avec la molécule qui doit réagir. Il s'agit d'une phosphorylation qui augmente le niveau d'énergie libre de cette molécule accepteuse, et lui permet de réagir avec d'autres molécules.
Si X + Y => Z - 2 kcal (réaction endergonique impossible spontanément)
Alors X + ATP => Xphosphate + ADP + 3 kcal (réaction exergonique)
Xphosphate + Y => Z + phosphate + 2,3 kcal (réaction exergonique)
Sachant que l'hydrolyse de l'ATP
donne => ADP + Pi + 7,3 kcal, la phosphorylation de la molécule X lui a fait
accroître son énergie libre de 4,3 kcal ( 7,3 - 3 = 4,3), la rend moins
stable, et de ce fait plus réactive que la molécule X initiale.
Le phosphate inorganique libéré par
l'hydrolyse de l'ATP ne peut pas se combiner avec une autre molécule d'ADP car
la répulsion électrostatique des ions phosphates entre eux les empêche de
reformer de l'ATP sans apport d'énergie extérieur.
Des phosphagènes (phosphocréatine
et phosphoarginine dans le muscle) permettent de restaurer rapidement l'ATP à
partir de l'ADP.
LA PHOTOSYNTHESE
1) CAPTURE DE L'ENERGIE SOLAIRE
Cette capture se fait grâce à la
chlorophylle et aux caroténoïdes, présents dans les plantes. La chlorophylle
est une grosse protéine s'organisant autour d'un noyau tétra pyrrolique :
l'hème qui renferme un atome de magnésium.
L'hémoglobine et la myoglobine
contiennent un hème qui renferme un atome de fer.
D'un point de vue physique, la configuration atomique de l'atome de magnésium est : 1s2 , 2s2, 2p6, 3s2, avec toutes ses orbitales saturées, alors que celle du fer est 1s2, 2s2, 2p6, 3s2, 3p6, 3d6, 4s2, avec l'orbitale 3d insaturée. Cela signifie que le niveau énergétique du dernier électron est moins élevé pour le magnésium que pour le fer. De ce fait le magnésium est plus réactif que le fer.
La lumière rencontrant la matière est réfléchie, absorbée ou transmise. Lorsqu'elle rencontre une feuille végétale, les radiations rouges et les bleues sont absorbées, les radiations vertes sont réfléchies ou transmises, c'est pourquoi les feuilles nous apparaissent vertes.
Chaque cellule végétale de la
feuille contient des chloroplastes. Ce sont des organites limités par une
double membrane externe contenant un liquide dense : le stroma, dans lequel sont
empilés des grana de thylakoïdes (sorte de poche). La membrane de ces
thylakoïdes contient des amas de centaine de molécules de chlorophylles et de
caroténoïdes.
Ces molécules de pigments sont
rangées en antennes de captation qui convergent vers un centre réactionnel.
Sous l'effet de la lumière, une
molécule de chlorophylle de ces antennes de captation absorbe un photon, ce qui
a pour effet de faire passer l'électron de son état fondamental stable à une
orbitale d'un niveau énergétique potentiel supérieur dans un état excité
instable.
Cette instabilité se transmet de
proche en proche lorsqu'il revient à son état fondamental, jusqu'à ce que
l'électron excité soit celui de la chlorophylle du centre réactionnel.
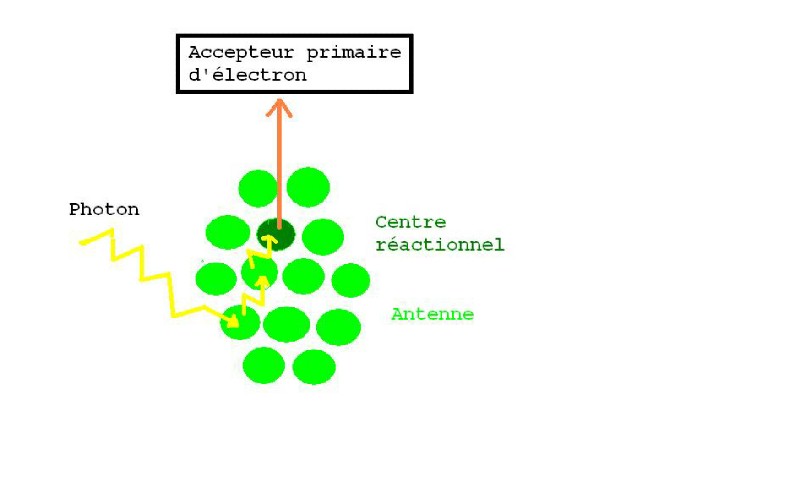
Un accepteur primaire d'électron se trouve près du centre réactionnel et va assurer le transport de l'électron en lui faisant perdre son énergie potentielle avant de le rendre à la chlorophylle du centre réactionnel dans son état fondamental. C'est un transport cyclique. Le transport non cyclique fait intervenir de l'eau dont l'hydrolyse rend son électron au centre réactionnel.
- Transport cyclique d'électron :
Il se fait à partir d'un centre réactionnel composé de deux molécules de chlorophylle a, appelées P700, qui absorbent entre autre la lumière ayant une longueur d'onde de 700 nm. C'est le photosystème I.
L'accepteur primaire capte l'électron excité du centre réactionnel P700, et le transmet à une protéine : la ferrédoxine, puis à une chaîne de cytochromes qui font descendre progressivement l'énergie potentielle de l'électron excité en libérant des protons (H+) dans l'espace intra-thylakoïdien. Ceux ci vont créer un gradient de H+ entre le stroma et l'espace intra-thylokoïdien. Le flux de H+ va rejoindre le stroma en passant dans une protéine membranaire tunnelisée : l'ATPsynthétase, qui va synthétiser de l'ATP.
- Transport non cyclique d'électron :
Il fait appel à un centre
réactionnel composé de deux molécules de chlorophylle a, appelées P680.
C'est le photosystème II.
Les ondes absorbées de 680 nm sont
plus énergétiques que pour la P700 car la loi de Planck nous indique que
l'énergie d'un rayonnement est inversement proportionnel à sa longueur d'onde
: E = h /
.
La lumière agit d'abord comme dans
le photosystème I. L'accepteur primaire capte l'électron excité du centre
réactionnel P700, et le transmet à la ferrédoxine, puis il est mis en
réserve dans le complexe NADPH + H+, selon la réaction
NADP+ + H2O + 2 e
=> NADPH + H+ + ½ O2
La chlorophylle P700 ayant perdu
un électron, devient un agent oxydant très puissant.
Le NADP+ est le nicotinamide adénine
dinucléotide phosphate
Les antennes du photosystème II
absorbent des photons, ce qui excite la chlorophylle P680 du centre
réactionnel, et lui fait perdre deux électrons qui sont pris par un accepteur
primaire qui les transmet à un complexe cytochrome. Pendant ce transport, les
électrons perdent de l'énergie potentielle jusqu'à atteindre P700 et comble
le vide laissé par la perte de deux électron au cours du photosystème I.
Il manque donc deux électrons au
photosystème II, sur la chlorophylle P680. Ils vont être fournis par une
hydrolyse de l'eau :
H2O => 2H+ + 2 e + ½ O2
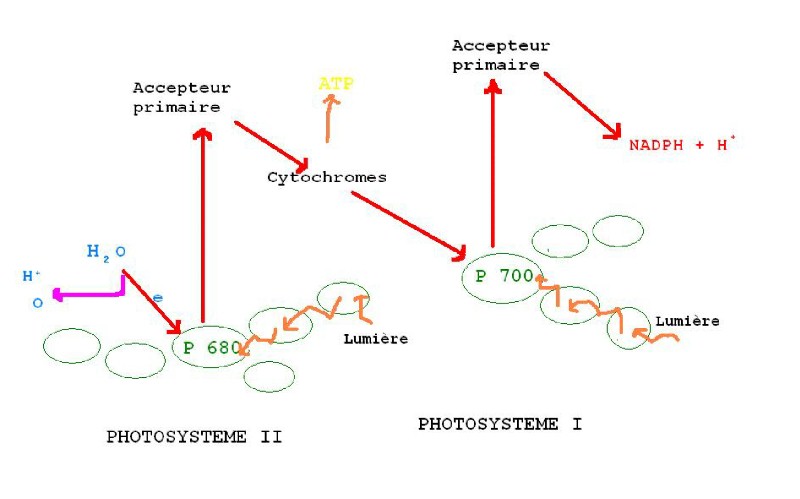
La baisse d'énergie potentielle à chaque transfert de cytochrome s'accompagne de l'émission de H+ à travers la membrane thylakoïde. Le gradient H+ obtenu entre l'espace inter membranaire thylakoïdien et le stroma, va amorcer la synthèse d'ATP à travers l'ATPsynthétase.
Au cours du photosystème II la chlorophylle P700 a perdu deux électrons qui ont été comblés par l'apport de deux électrons de la chlorophylle P680 du photosystème I. La P680 retrouve deux électrons suite à la scission enzymatique de l'eau, et il y a libération accessoire d'oxygène.
La lumière engendre un courant
d'électrons et de protons qui produisent de l'ATP et du NADPH + H+ .
La chlorophylle a donc converti de l'énergie lumineuse en énergie chimique, en
consommant de l'eau et en faisant apparaître de l'oxygène qui n'est qu'un sous
produit..
2) SYNTHESE DES MATIERES ORGANIQUES
La synthèse des matières organiques
se fait le jour, au cours du cycle de Calvin pendant lequel du dioxyde de
carbone est transformé en glucide. Ces réactions ne sont possibles que grâce
à la présence d'ATP pour fournir de l'énergie et du NADPH + H+ pour céder
des électrons.
La ribulose diphosphate
carboxylase catalyse la fixation du CO2 sur le ribulose diphosphate (RuDP). Cela
donne un composé en C6 très instable, qui se scinde aussitôt en deux
molécules en C3. La phosphorylation de ces molécules avec de l'ATP augmente
leur énergie potentielle. Puis une paire d'électron est cédée par NADPH + H+
tandis qu'un phosphate inorganique est cédé. On obtient ainsi du
phosphoglycéraldhyde (PGAL) qui va être la base de toutes les synthèses de
matière organique : glucides, lipides, protides.
Pour finir, 5 moles de PGAL avec 3 moles d'ATP vont reconstituer 3 moles de RuDP
pour réamorcer le cycle.
En résumé, dans une cellule végétale ont lieu des réactions biochimiques dans les chloroplastes où l'énergie solaire est transformée en énergie chimique puis le cycle de Calvin synthétise tout ce dont la plante a besoin pour se développer.
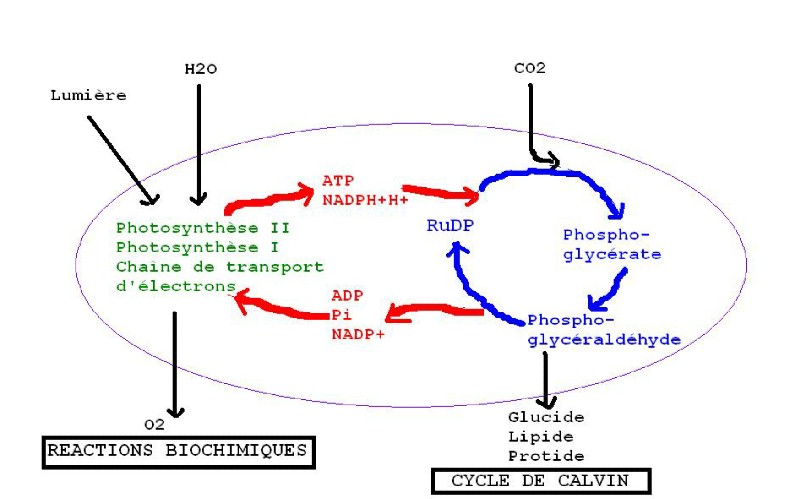
TRANSFORMATION DU VEGETAL EN ATP
L'herbivore et le cheval en
particulier, consomme les végétaux. Son système digestif va dégrader ces
aliments pour les réduire à l'état de glucose, d'acide gras et d'acides
aminés aptes à passer dans les cellules.
Le glucose dont la synthèse provient de l'absorption de radiations solaires contient donc de l'énergie et il correspond à une diminution de l'entropie du système (degré de désorganisation de la matière).
La combustion du glucose avec de l'oxygène libère 686 kcal/mole
C6 H12 O6 + 6 O2 => 6 CO2 + 6 H2O + 686 kcal
Cette énergie correspond à la restitution de l'énergie de rayonnement qui avait été utilisée par la plante pour synthétiser le glucose. Elle est donc contenue dans chaque molécule de glucose.
Lors de la respiration cellulaire
toute cette énergie ne va pas être dissipée en chaleur, une partie va être
stockée sur des atomes hautement énergétiques, en particulier le phosphore.
En effet, l'adénosine triphosphate
ATP se termine par trois phosphates, chaque phosphate PO3 - -
est en répulsion avec son voisin, et cette énergie de répulsion vaut 7,3 kcal
/ mole lorsque ATP => ADP + P . C'est la valeur de l'énergie libre de l'ATP.
L'oxydation du glucose se fait par
échange d'électron de la molécule donneuse dite " réducteur " (ici
le glucose) vers la molécule receveuse dite " oxydante " (ici
l'oxygène). Ceci forme un couple redox dans lequel les électrons se déplacent
vers un état plus stable en diminuant leur énergie potentielle mais qui
s'accompagne d'une augmentation de l'entropie du système.
Cette oxydation totale du glucose en
CO2 + H2O en une seule réaction libérant 686 kcal sous forme de chaleur n'est
pas utilisable par la cellule vivante. Aussi le métabolisme d'une cellule
aérobie va utiliser des composés intermédiaires contenant encore de
l'énergie, donc encore réactifs, avant d'amener les électrons à leur plus
bas niveau d'énergie.
Ces composés intermédiaires ont un
potentiel réducteur moindre car possédant moins d'électrons libres captables
par l'oxygène.
Les réactions d'oxydation sont donc
des réactions de transfert d'électrons.
La respiration cellulaire (oxydation du glucose) se fait en trois phases :
- la glycolyse dans le cytosol transforme le glucose en pyruvate et en ATP
- le cycle de Krebs dans la mitochondrie transforme le pyuvate en CO2 en ATP, en
NADH + H+ et en FADH2
- la chaîne de transport d'électrons dans la mitochondrie transforme NADH + H+
, FADH2 en ATP
1) LA CHAÎNE RESPIRATOIRE
Dans les mitochondries des cellules, le transfert d'électrons est réalisé
par des enzymes : les déshydrogénases qui captent des électrons avec des
protons.
Il y a la nicotinamide adénine dinucléotide NAD+ et la flavine adénine
dinucléotide FAD qui agissent ainsi :
Substrat + NAD+ => substrat oxydé + ( NADH + H+) réduit
Substrat + FAD => substrat oxydé + ( FADH2 ) réduit
La forme réduite de FAD qui est FADH2, renferme une grande quantité d'énergie libre. Ainsi, l'oxydation de FADH2 par l'oxygène libère 52 kcal / mole
FADH2 + ½ O2 => FAD + H2O + 52 kcal
Cependant, dans la cellule, ces enzymes ( NAD+ et FAD) ne peuvent pas être directement oxydées par l'oxygène, la libération brutale de 52 kcal que représente leur énergie chimique potentielle, pourrait endommager la cellule, et cette énergie serait perdue. Une chaîne respiratoire de 7 transporteurs d'électrons, rechargeant l'ADP en ATP, va permettre aux électrons d'augmenter leur entropie par étape jusqu'à leur réaction finale avec l'oxygène. Chacun des transporteurs d'électrons successifs a une pression électronique (faculté de perdre cet électron) plus faible que son prédécesseur. On obtient une chaîne respiratoire dans laquelle chaque transporteur a une place bien définie, fonction de sa pression électronique.
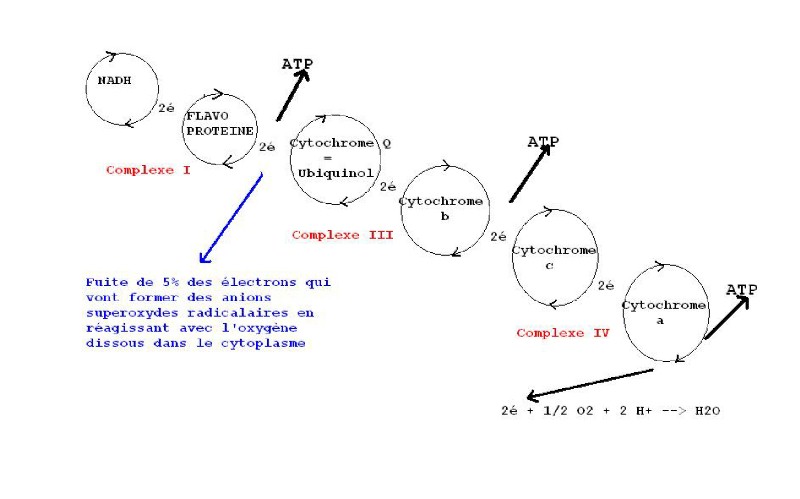
Les cytochromes sont des protéines organisées autour de l'hème contenant un atome de fer qui est oxydé ou réduit pendant le transport. L'hème des cytochromes est de même nature que celui de l'hémoglobine et de la myoglobine, et il subit les mêmes dégradations métaboliques vers la bilirubine (voir suivi biologique du cheval d'attelage).
A chaque transfert de transporteur, il y a une petite libération d'énergie qui accompagne la montée d'entropie de l'électron et qui est suffisante pour synthétiser l'ATP. Cela se fait lors du transfert de Flavoprotéine vers Coenzyme Q, Cytochrome b vers c, Cytochrome a vers O2.
C'est la pression en O2, accepteur final d'électron, qui entraîne le déroulement de cette chaîne de transport et libère de l'ATP. La chaîne respiratoire est aérobie stricte.
2) LA GLYCOLYSE
Elle a lieu dans le cytosol de toute cellule. Dans un premier temps une
enzyme : l'hexokinase augmente l'énergie libre du glucose en le phosphorylant,
avec la consommation d'une mole d'ATP. Après une isomérisation en fructose
6-phosphate et une nouvelle phosphorylation, le fructose 1,6-diphosphate est peu
stable et se coupe en deux trioses phosphates.
Le phosphoglycéraldéhyde subit une déshydrogénation, incorpore un phosphate
inorganique, puis cède deux ATP pour former du pyruvate. L'autre triose se
transforme en phosphoglycéraldéhyde grâce à une isomérase et va finir aussi
en pyruvate.
Au total, à partir d'une mole de glucose, la glycolyse donne deux moles de pyruvate + 2 ATP + 2 NADH + H+
La glycolyse ne nécessite pas la présence d'oxygène, et ne libère pas de dioxyde de carbone. Elle libère moins du quart de l'énergie contenue dans le glucose.
3) LE CYCLE DE KREBS
En présence d'oxygène le pyruvate quitte le cytosol pour pénétrer dans la mitochondrie. Là il subit une décarboxylation et une oxydation pour devenir un acétate. Il s'unit alors au CoA par un pont de soufre instable, ce qui rend le groupement acétyl très réactif.
L'acétyl-CoA libère son acétate dans le cycle de Krebs dont les 8 étapes finissent par donner 3 moles de NADH + H+, 1 mole de FADH2, 1 mole d'ATP, 2 moles de CO2
Dans la mitochondrie la chaîne de transport d'électron assure le passage des protons de la matrice vers l'espace inter membranaire des crêtes à chaque transfert d'électron, par une réaction exergonique. Ainsi la concentration en H+ de l'espace inter membranaire peut devenir 100 fois plus grande que celle de la matrice. Ce gradient de protons est à l'origine de leur retour vers la matrice, à travers un canal formé dans l'ATPsynthétase (protéine structurelle de la membrane mitochondriale ). Ce passage de H+ va produire une phosphorylation de l'ADP, et donner de l'ATP.
Au total 1 mole de glucose + 6 O2 donne 36 ATP, 6 CO2 , 6 H2O au cours de la glycolyse, du cycle de Krebs et de la chaîne de transport d'électron.
La respiration cellulaire aérobie a un rendement de 63% ( la voiture convertit l'énergie libre de l'essence en énergie cinétique avec un rendement de 25%) . Les 37% d'énergie dissipée en chaleur sont utilisés pour maintenir la température du cheval à 37° - 37°5
Sans oxygène, la phosphorylation oxydative cesse car la chaîne respiratoire
s'arrête faute de couplage de l'électron du cytochrome a3 avec ½ O2 +
H+ pour
donner H2O
On peut obtenir une fermentation alcoolique lorsqu'à la fin de la glycolyse, le
pyruvate libère du CO2 et devient de l'éthanol, ou une fermentation lactique
lorsque le pyruvate se fait réduire directement par NADH + H+ et devient du
lactate.
ATP et ENERGIE CINETIQUE
Chez le cheval, l'énergie chimique de l'ATP obtenue par oxydation des aliments, va être transformée en énergie cinétique par l'intermédiaire des muscles qui vont consommer l'ATP.
1) LE MUSCLE SQUELETTIQUE
Le muscle squelettique est constitué de faisceaux de fibres musculaires de 0,1 à 1 mm de diamètre et de grande longueur. Chaque fibre musculaire d'un diamètre de 0,01 à 0,1 mm est une cellule.
La fibre musculaire est donc limitée par une membrane cellulaire (le sarcolemme) , et renferme des centaines de myofibrilles baignant dans le sarcoplasme qui contient tous les organites de vie de la cellule.
Quelques particularités :
- la fibre musculaire contient plusieurs noyaux cellulaires
- elle contient une grande quantité de mitochondries propres à synthétiser de
l'ATP en aérobiose ( l'entraînement du cheval à l'endurance fait accroître
son pool de mitochondries)
- elle contient une grande quantité de vacuoles remplies de lipides, des
inclusions de glycogène
- son réticulum endoplasmique (RE) est formé de compartiments fermés
disposés en tubules longitudinaux parallèles aux myofibrilles. Ces tubules
sont des réservoirs de Ca++. La membrane cellulaire présente des invaginations
profondes, transversales aux myofibrilles en de nombreux endroits. Cela forme
les tubules transverses appelés système T, qui sont en contact étroit avec
les vésicules terminales des tubules longitudinaux du RE. Cet ensemble a une
grande importance car il est à l'origine du déclenchement de la contraction
musculaire.
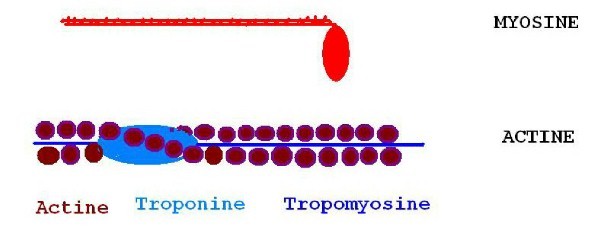
- Les myofibrilles font l'originalité de la cellule musculaire. Chaque
myofibrille est constituée d'une succession de plusieurs fois 2000 filaments
fins d'actine et de 1000 filaments épais de myosine. C'est le mouvement relatif
de la myosine sur l'actine qui aboutit à une contraction du muscle.
Le filament d'actine est constitué de deux brins torsadés d'une protéine globulaire (la G actine) qui a été polymérisée. Des molécules filamenteuses de tropomyosine parcourent le cœur de la torsade, et à intervalles réguliers, des protéines globulaires (la troponine) sont attachées au filament de tropomyosine.
La myosine est constituée de deux chaînes protéiques lourdes, torsadées se terminant à une extrémité par une double tête globulaire qui renferme aussi des chaînes légères sièges de la fixation du Ca++.
2) CONTRACTION MUSCULAIRE
Elle se fait par glissement de la myosine sur l'actine , en présence de Ca++, Mg++, ATP, ATPase.
La contraction est déclenchée par l'émission d'un potentiel d'action (PA)
du motoneurone de la moelle épinière. Ce PA arrive à la terminaison nerveuse
de la plaque motrice (jonction entre le nerf et le muscle) où il provoque
l'apparition d'un courant entrant de Ca++ de l'espace synaptique vers la
terminaison nerveuse. Ceci entraîne la libération massive d'acétylcholine
contenue dans des vésicules de la zone pré synaptique vers l'espace
synaptique.
Côté post synaptique, des récepteurs d'acétylcholine vont être stimulés et
vont déclencher une réponse sous forme d'un potentiel d'action sur le
sarcolemme. L'acétylcholine est alors hydrolysée par l'acétylcholinestérase,
ce qui rend possible la repolarisation de la plaque motrice et la laisse dans
l'attente d'un nouveau PA.
Le PA musculaire se propage au niveau du système T de toutes les fibres musculaires et provoque la libération du Ca++ des tubules longitudinaux. Ceci multiplie par 1000 la concentration en Ca++ dans les fibres musculaires. Le Ca++ va se fixer sur la troponine et initialiser la réaction actine-myosine.
Au repos un bouchon, en contact avec un récepteur du tubule T, ferme un canal à Ca++ du RE (réticulum endoplasmique). Au passage du PA, le bouchon se déplace, ce qui libère le Ca++ vers le sarcoplasme d'où il se lie à la troponine. Quand le potentiel de membrane revient au repos, le bouchon reprend sa place et les pompes à Ca++ de la membrane du RE font revenir le Ca++ dans le RE en consommant de l'ATP. Il faut 2 ATP pour pomper 1 Ca++ . Le Ca++ est stocké dans le RE lié à une protéine (la calséquestrine). Ainsi séquestré, le Ca++ ne présente pas de gradient de concentration nécessitant une activité de pompage permanente, consommatrice d'ATP.
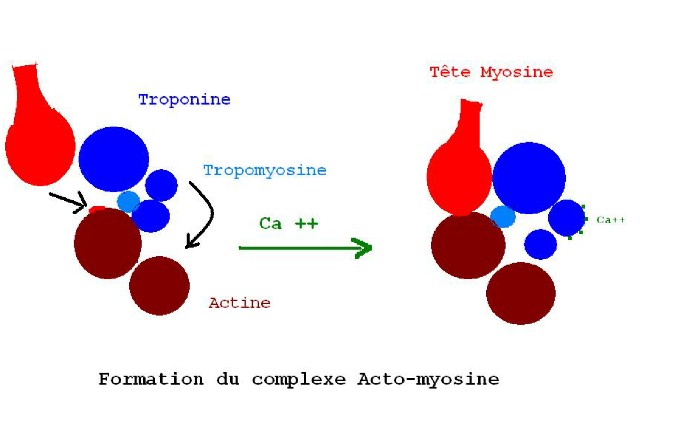
RÉACTION ACTINE - MYOSINE
Le Ca++ se fixe sur la troponine dont la structure globulaire subit alors un
changement de conformation spatiale, dégageant sur l'actine le site de fixation
de la tête de la myosine. Chaque complexe de troponine fixe 4 ions Ca++ .
Les deux têtes de la myosine ont été réunies en une tête avec de l'ATP pour
former un complexe ATP-myosine
Le complexe ATP-myosine créé un pont avec l'actine et aussitôt l'ATPase en présence de Mg++ va hydrolyser l'ATP du complexe ATP-myosine-actine, avec dégagement de chaleur car c'est une réaction exothermique. La perte du Phosphate inorganique modifie l'organisation spatiale du complexe ADP-myosine-actine. La tête de myosine qui faisait un angle de 90° avec sa queue, ne fait plus qu'un angle de 50° après la perte du Pi, puis un angle de 45° après la perte de l'ADP. Ce mouvement angulaire entraîne le glissement de l'actine sous la myosine.
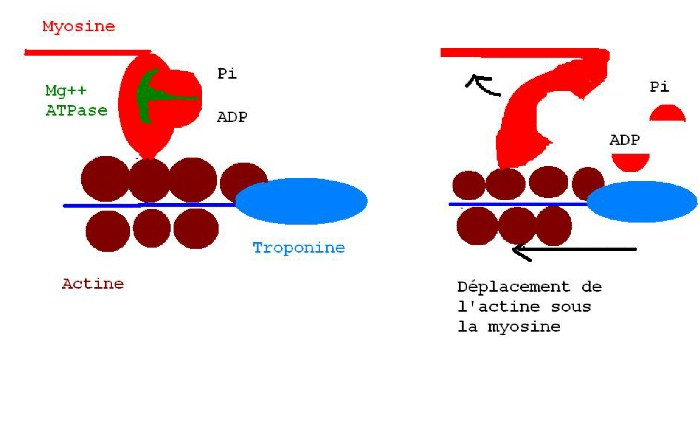
Lorsque la concentration en ATP sera suffisante, les ponts entre la myosine et l'actine disparaîtront, et en même temps les deux têtes de la myosine se redresseront à 90°, puis l'ATP se fixera à nouveau sur la myosine pour former le complexe ATPmyosine . Ainsi la fibre retrouvera son état initial de relâchement, prête à subir un nouveau PA
L'absence de synthèse de l'ATP sur un organisme mort laisse persister les
ponts actine-myosine et fait cesser le pompage du Ca++ vers les tubules T . Le
muscle devient inextensible, c'est l'état de rigidité cadavérique qui va
persister tant que l'actine et la myosine ne seront pas décomposées.
ÉNERGÉTIQUE DE LA LOCOMOTION
La concentration en ATP est presque la même dans le muscle au repos et dans
le muscle contracté. Cela indique qu'une autre source d'ATP s'ajoute à l'ATP
libre du sarcoplasme. Il s'agit d'un phosphagène qui génère rapidement de l'ATP.
Le principal est la phosphocréatine qui cède son phosphate à l'ADP sous
l'effet de la créatine phosphokinase :
Phosphocréatine + ADP => créatine + ATP
La concentration en phosphocréatine des fibres musculaires est bien plus élevée que la réserve d'ATP. Lorsque le métabolisme oxydatif (aérobie) est en pleine activité, les synthèses d'ATP qu'il produit suffisent à entretenir l'activité musculaire. Les phosphagènes interviennent quelques secondes après le début du mouvement, lorsque l'ATP libre a été consommé.
Plus la vitesse de recyclage des ponts acto-myosine est élevée, plus la dépense énergétique du raccourcissement musculaire sur une distance parcourue donnée est élevée. Ceci est du à une mise en œuvre plus intensive des pompes à Ca++. C'est ainsi que le cheval trouve sa meilleure allure (pas, trot ou galop) pour une vitesse demandée, avec un effort de traction donné, qui lui permet de consommer le moins d'énergie possible.
Pour une vitesse donnée, la dépense énergétique sera plus faible pour les gros animaux (cheval), que les moyens (chien) et que les petits (rat, souris). Cela est la conséquence de la vitesse de contraction musculaire qui est plus grande chez les petits animaux, et qui nécessite donc une plus grande action des pompes à Ca++ , consommatrices d'ATP.
Le coût énergétique du transport par kg et par mètre est
- élevé à faible vitesse car la durée du transport est longue
- faible à vitesse intermédiaire
- élevé à haute vitesse car la vitesse des contractions musculaires augmente
et l'utilisation des pompes à Ca++ aussi. (voir " l'alimentation du cheval
d'attelage ")
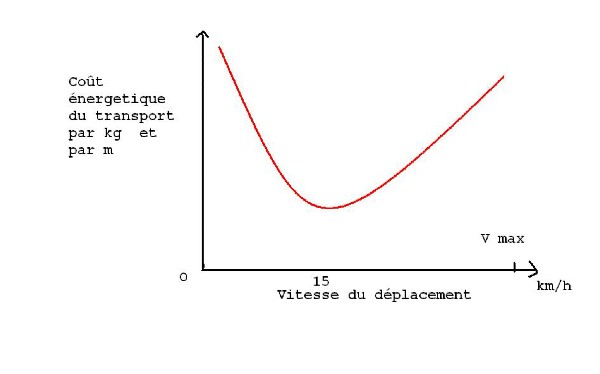
Pour un cheval la vitesse optimum de déplacement est d'environ 15 km/h, elle
est de 12 km/h pour un poney moyen ou un cheval de trait. Ce sont d'ailleurs les
vitesses qui avaient été retenues lors de l'élaboration des règlements des
concours d'attelage pour la maniabilité et les phases A et E du marathon.
Dr Guy SOUFFLEUX
BIBLIOGRAPHIE
CAMPBELL . " Biologie ". Ed De Boeck Université, Quebec 1995, 1254 pages
ECKERT Roger. " Physiologie animale ". Ed De Boeck Université , Paris 1999, 822 pages
HECHT Eugène. " Physique ". Ed De Boeck Université, Paris 1999, 1304 pages
SILBERNAGL S. DESPOPOULOS A. " Atlas de poche de physiologie ". Ed
Flammarion. Paris 1993, 366 pages
MOTS CLES
Cheval - photosynthèse - énergie musculaire - contraction musculaire
Retour à la page d'accueil AVEMA